| Aged skin = شيخوخة الجلد |
|
|
Aging of Skin
▪ EFFECTS OF AGING In both developed and developing nations, the number and proportion of older people are increasing. Thirty-one percent of the U.S. population is expected to be 55 years of age or older by the year 2030.1 This demographic shift compels health care providers and government officials to confront the pathophysiology of aging and associated health issues. Aging is a process of progressive decreases in the maximal functioning and reserve capacity of all organs in the body, including the skin. This naturally occurring functional decline in the skin is often compounded and accelerated by chronic environmental insults, such as ultraviolet (UV) irradiation. Aging occurs at the cellular level and reflects both a genetic program and cumulative environmentally imposed damage. Mammalian cells can undergo only a limited number of cell divisions and then arrest irreversibly2-4 in a state known as replicative senescence, after which they are refractory to mitogenic stimuli. This fact has led to the perception that aging evolved in multicellular organisms as a cancer prevention mechanism because it prevents the unlimited and possibly unregulated growth of cells whose DNA has been progressively damaged over their life span. Of note, in general the more proficient the DNA repair mechanisms of the organism, the longer its life span. Furthermore, there is an inverse correlation between the organism's life span and metabolic rate consistent with the understood role of cumulative oxidative DNA damage, due to aerobic metabolism, in the aging process.
▪ AGING MECHANISMS Telomeres and Aging Telomeres, the terminal portions of eukaryotic chromosomes, consist of up to many hundreds of tandem short sequence repeats (TTAGGG in all mammals). During mitosis of somatic cells, DNA polymerase cannot replicate the final base pairs of each chromosome, resulting in progressive shortening with each round of cell division. A special reverse transcriptase, telomerase, can replicate these chromosomal ends, but, with the exception of stem cells and germline cells, the enzyme is normally expressed at extremely low levels, insufficient to maintain telomere length. Human telomeres shorten more than 30 percent during adulthood even in relatively quiescent skin fibroblasts, and telomeres of patients with premature aging syndromes, such as Werner syndrome progeria,and dyskeratosis congenita, are shorter than those of age-matched controls. Critically short telomeres signal for proliferative senescence or apoptosis, depending on cell type, and appear to compromise DNA stability and transcription of subtelomeric genes,12 presumably contributing to the aged phenotype. Thus, telomeres appear to serve as a biologic clock that determines proliferative life span of the cell.
DNA Damage and Aging In humans, the only genes implicated in the rate of aging are those in which mutations are responsible for premature aging syndromes. For example, Cockayne syndrome patients display mutations in DNA helicases, enzymes that participate in the repair of DNA damageataxia telangiectasia is caused by a mutation in the ATM gene, encoding a kinase that senses DNA damage; and Werner syndrome is caused by mutation in a protein with DNA helicase and exonuclease domains.Progeria, a disease that leads to death from atherosclerotic heart disease often in the first decade of life, is caused by mutations in lamin A, a protein important in maintaining chromatin organization in the nucleus for transcriptional control and for DNA damage repair.17,18 These human premature aging diseases suggest that decreased DNA repair capacity is associated with accelerated aging and that cumulative DNA damage plays a major role in the aging process. Still, the role of these genes in normal aging is not established, as patients with so-called premature aging syndromes display some manifestations of aging at an accelerated rate but lack other features of normal aging and have characteristic findings that differ greatly from those of normal aging. Aging and the Immune System The immune system has two major roles: defense against external insults and internal immunologic surveillance. The function of antigen-presenting cells, antigen-specific B and T cells, and lymphocyte cytokine secretion declines with age. Conversely, the level of pro-inflammatory cytokines increases with age. These changes have been attributed to oxidative stress and contribute to the increased incidence of infections and malignancies in the elderly. ▪ SKIN AGING Cutaneous aging includes two distinct phenomena: Intrinsic aging is a universal, presumably inevitable change attributable to the passage of time alone; photoaging is the superposition on intrinsic aging of changes attributable to chronic sun exposure, which are neither universal nor inevitable. The former is
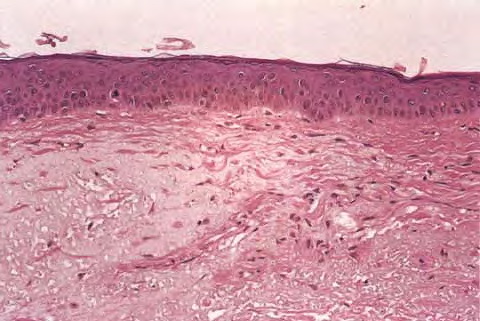
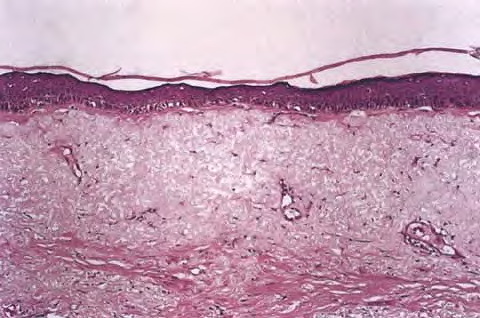
Intrinsic Skin Aging The skin changes that occur with aging (Table 108-1) lead to a gradual physiologic decline (Table 108-2).21 Major age-related changes in the skin's appearance include dryness (roughness), wrinkling, laxity, and a variety of benign neoplasms. Aged skin is inelastic and recovers more slowly after injury. MECHANISMS OF INTRINSIC SKIN AGING A major aging theory suggests that cumulative damage to biomolecules, including DNA as a result of continuous generation of free radicals, results in increased cellular vulnerability and eventually terminates in senescence or apoptosis. The skin, like other bodily systems, is continuously exposed to reactive oxygen species (ROS) generated during aerobic metabolism . Although the skin contains a network of antioxidant enzymes (superoxide dismutases, catalase and glutathione peroxidase) and nonenzymatic anti-oxidant molecules (vitamin E, coenzyme Q10, ascorbate, and carotenoids), this system is less than completely effective and tends to deteriorate with aging.
Oxidative stress upregulates the level of stress regulatory proteins, one of which is nuclear factor κB, a transcription factor that induces the expression of pro-inflammatory cytokines like interleukin (IL)-1 and IL-6, vascular endothelial growth factor (VEGF), and tumor necrosis factor (TNF)-α. These proteins are involved in immunoregulation and cell survival stimulate the expression of matrix-degrading metalloproteins,25 and are believed to play a central role in the aging process. Oxidative damage also affects telomeres. A recent hypothesis suggests a common cellular signaling pathway activated by DNA damage and involving the terminal portion of the telomeres. The terminal portion of the 3′ telomeric strand extends beyond the complementary 5′ strand (Fig. 108-1B), leaving a single stranded G-rich overhang. It is suggested that during both telomere shortening and repair of telomere damage, such as that encountered by oxidative stress, the normal loop structure at the end of telomeres is disrupted, exposing the 3′ overhang that under baseline conditions is “buried” in the loop structure. Exposure of the TTAGGG tandem repeat sequence then appears to activate p53 and to stimulate p53-dependent responses known to include proliferative senescence and apoptosis. Thus, the intrinsic component of skin aging involves progressive oxidative stress and telomere signaling as telomeres shorten during serial cell division and in response to oxidative DNA damage. EPIDERMIS The most striking and consistent histologic change is flattening of the dermal-epidermal junction with effacement of both the dermal papillae and epidermal rete pegs. This results in a considerably smaller surface between the epidermis and dermis and presumably less communication and nutrient transfer. Dermal-epidermal separation has been demonstrated to occur more readily in old skin, undoubtedly explaining the propensity of the elderly to torn skin and superficial abrasions after minor trauma. There is an age-associated epidermal thinning of 10 percent to 50 percent between the ages of 30 and 80 years. Variability in epidermal thickness and individual keratinocyte size increases, including those of the basal layer. Evidence suggests that epidermal keratinocytes senesce, and senescent cells are more resistant to apoptosis. Such keratinocytes are therefore more likely to accumulate mutations, increasing their risk for malignant transformation. Studies also show loss of epidermal stem cell population in aged skin as determined by the loss of cells expressing CD71 (transferrin receptor) and α6 integrin, accepted markers for keratinocyte stem cells.37 At the electron microscopic level, sun-protected old skin is characterized by some widening of inter-keratinocyte spaces, by reduplication of the lamina densa and anchoring fibril complex in the basement membrane zone, and by loss of the numerous microvillous projections of basal cell cytoplasm into the dermis.36 Average thickness and degree of compaction of the stratum corneum appear constant with increasing age, although individual corneocytes become larger. The skin surface pattern, a patchwork of fine lines possibly determined by papillary dermal architecture, reveals slight age-associated loss of regularity. Age effects on percutaneous absorption depend in part on drug structure, with hydrophilic substances such as hydrocortisone and benzoic acid being less well absorbed through the skin of old versus young individuals but with hydrophobic substances such as testosterone and estradiol being equally well absorbed. Of perhaps greater clinical importance, aging markedly delays the recovery of barrier function in damaged stratum corneum, apparently because of slow replacement of neutral lipids, leading to decreased amount of lipids in the newly formed lamellar bodies.39 Lipid synthesis and activities of enzymes required to generate stratum corneum lipids decrease with age possibly because of aberrations in elements that regulate enzyme transcription, or abnormal autocrine/paracrine signaling.
In the elderly, the skin often appears dry and flaky, especially over the lower extremities, an area in which a remarkable age-associated decrease in the content of epidermal filaggrin has been reported.41 Filaggrin, required for binding of keratin filaments into macrofibrils, is also decreased in the skin of patients with ichthyosis vulgaris, and its lack has been postulated to cause the increased scaliness in both conditions.41 Barrier function also may be affected by this structural change. Epidermal turnover rate and thymidine-labeling index decrease approximately 30 percent to 50 percent between the third and eighth decades, with a corresponding prolongation in stratum corneum replacement rate. Linear growth rates also decrease for hair and nails. Epidermal repair rate after wounding likewise declines with age. A decrease in the number of enzymatically active melanocytes per unit surface area of the skin, approximately 10 percent to 20 percent of the remaining cell population each decade, has
Between early and late adulthood there is a 20 percent to 50 percent reduction in the number of morphologically identifiable epidermal Langerhans cells, the skin's immune effector cells responsible for antigen presentation. The remaining cells display morphologic abnormalities, including less and shorter dendrites, and they display reduced antigen-presenting capacity. These changes, compounded by decreases in cytokine production by keratinocytes and lymphocytes and failure of migration through the lymphatic system, presumably contribute to the observed age-associated decrease in cutaneous immune responsiveness. An endocrine function of human epidermis that declines with age is vitamin D production. Vitamin D, by binding its nuclear receptor, induces the transcription of numerous genes. Vitamin D deficiency leads to osteoporosis and recently has been implicated also in diabetes, hypertension, and prevalent tumors.45 Elderly individuals frequently have reduced serum levels of vitamin D, leading to osteomalacia, the decreased mineralization of bone classically associated with vitamin D deficiency. Although avoidance of dairy products (the principal dietary source of vitamin D), insufficient sun exposure, and sunscreen use undoubtedly contribute to vitamin D deficiency in the elderly, the level of epidermal 7-dehydrocholesterol per unit skin surface area also appears to decrease linearly by approximately 75 percent between early and late adulthood, suggesting that lack of its immediate biosynthetic precursor also may limit vitamin D production. Together these observations argue for use of vitamin D dietary supplements in the elderly. With regard to susceptibility to oxidative damage, there is progressive accumulation of damaged cellular proteins and lipids with aging. Furthermore, antioxidant defense systems decline with age, and, in addition, there is a decrease in DNA damage repair capacity.36 These changes in combination increase cellular mutability or their tendency to become senescent, or both. DERMIS Loss of dermal thickness approaches 20 percent in elderly individuals, although in sun-protected sites significant thinning occurs only after the eighth decade. Old skin is relatively acellular and avascular, and there is age-related loss of normal elastic fibers and dermal collagen. Decreased inflammatory responses in the elderly are the result of decreased synthesis and secretion of keratinocyte-derived cytokines and inflammatory mediators in addition to decreased endothelial response. The dermal microvasculature in middle-aged or elderly subjects also may show mild vascular wall thickening, especially in the lower legs as a result of gravitational forces; vascular wall thinning to less than one-half the normal young adult measurement, associated with absent or reduced perivascular veil cells, has been reported in skin of very elderly subjects and probably contributes to vascular fragility. Loss of elastin contributes to vascular rigidity. Electron microscopic studies show focal degeneration of the elastic component of dermal arterioles. The striking age-associated loss of vascular bed, especially of the vertical capillary loops that occupy the dermal papillae in young skin, and increased distance from the epidermis of existing loops, is thought to underlie many of the physiologic alterations in old skin, including pallor, decreased skin temperature, and the approximately 60 percent reductions in basal and peak induced cutaneous blood flow. VEGF of epidermal origin appears to play a major role in maintaining dermal vasculature, inducing the expression of anti-apoptotic proteins in endothelial cells, and decreased VEGF level in aged skin probably contributes to endothelial cells apoptosis. Also, recent evidence suggests that there is an age-associated decline of both angiogenic and anti-angiogenic factors, disrupting cutaneous angiogenic homeostasis.54 Decreased endothelial cell permeability response and decreased capacity to induce white cell adhesion55 contribute to the compromised immune response. When exposed to intense heat or cold, aging vessels demonstrate reduced ability to constrict, dilate, or shunt. Compromised thermoregulation, which predisposes the elderly to sometimes fatal heat stroke or hypothermia, may be due in part to reduced vasoactivity of dermal arterioles and, in the latter instance, to loss of subcutaneous fat as well. Reduction in the vascular network surrounding hair bulbs and eccrine, apocrine, and sebaceous glands may contribute to their gradual atrophy and fibrosis with age. Age-associated decreases in wheal resorption and dermal clearance of transepidermally absorbed materials have been reported, probably due to alterations in both the vascular bed and the extracellular matrix. Conversely, the time required for development of a tense blister after topical ammonium hydroxide application is nearly twice as long in older individuals, suggesting a decreased transudation rate with age in injured skin. Impaired transfer of cells as well as solutes between the extravascular and intravascular dermal compartments is suggested by several studies; multiple factors undoubtedly contribute. With aging there is a decrease in the density and lumen size of lymphatic vessels accompanied by increased rigidity and decrements in lymphatic drainage, affected no doubt by decreased surrounding elastic fibers. Biochemical changes in collagen, elastin, and dermal ground substance lead to increased skin rigidity primarily due to modifications in collagen. Collagen content per unit area of skin surface decreases approximately 1 percent per year throughout adult life, and the remaining collagen fibrils appear disorganized, more compact, and granular, and they display increased collagen cross-links. The latter is the result of decreased collagen I and III synthesis; decrements in enzymatic processing of collagen as well as nonenzymatic glycosylation, a process that leads to molecular damage of proteins with a long half-life such as collagen36; and increased collagenase levels. Such changes almost certainly contribute to impaired wound healing in the elderly. Beginning in early adulthood, elastic fibers decrease in number and diameter; by old age, they often appear fragmented, with small cysts and lacunae, especially in the dermal-epidermal junction most likely due to enzymatic degradation of elastin. Elastic fibers also show progressive cross-linkage and calcification with age. At the biochemical level, there is an age-associated decrease in numerous elastic fiber components, including elastin, fibrillin, and fibulin-2. With aging, the level of fibulin-5, an extracellular matrix protein that functions as a scaffold for elastic fibers, appears to decrease before other changes are observed, suggesting that loss of fibulin-5 is a marker for skin aging. The ground substance mucopolysaccharides, glycosaminoglycans (GAGs), and proteoglycans are decreased relative
Changes with age in the mechanical properties of the skin during adulthood include progressive loss of elastic recovery, consistent with gradual destruction of the dermal elastic network, and marked prolongation of the time required for excised skin to return to its original thickness. In vivo ultrasound studies also show age-associated differences in water distribution in the dermis,66 no doubt affecting dermal pliability, resilience, and elasticity. Overall, a picture emerges of aging dermis as an increasingly rigid, inelastic, and unresponsive tissue that is less capable of undergoing modifications in response to injury or stress. SUBCUTANEOUS TISSUE, MUSCLES, AND BONE Like other striated muscles, facial muscles show accumulation of the “age pigment” lipofuscin, a marker of cellular damage. Compounded by diminished neuromuscular control, this deterioration contributes to wrinkle formation.67 In addition, subcutaneous fat is depleted from distinct facial regions, including the forehead, preorbital, buccal, temporal, and perioral regions. In contrast, there is a prominent increase in fatty tissue in other areas, including the submental regions, the jowls, the nasolabial folds, and the lateral malar areas. In contrast to the young face in which fat is diffusely dispersed, fat in the aged face, subject to the force of gravity, contributes to sagging and drooping of the skin. Finally, like other parts of the skeleton, facial bones display reduced mass with age. Bone resorption affects particularly the mandible, maxilla, and frontal bones. Bone loss in these areas enhances facial skin droopiness and contributes to the obliteration of the demarcation between the contour of the jaw and the neck that is so distinct in young adults. HAIR By the end of the fifth decade, approximately one-half the population has at least 50 percent gray (white) scalp hair, and virtually everyone has some degree of graying due to progressive and eventually total loss of melanocytes from the hair bulb.70 Loss of melanocytes is believed to occur more rapidly in hair than in skin because the cells proliferate and manufacture melanin at maximal rates during the anagen phase of the hair cycle, whereas epidermal melanocytes are comparatively inactive throughout their life span. More specifically, hair graying reflects loss of the melanocyte stem cell population in hair follicle bulge due, at least in part, to compromised interaction between two transcription factors, microphthalmia-associated transcription factor (Mitf) and Pax3. Scalp hair may gray more rapidly than other body hair because its anagen to telogen ratio (see Chap. 84) is considerably greater than that of other body hair. Advancing age is also accompanied by a modest decrease in number of hair follicles, due in part to atrophy and fibrosis. In addition, with aging there is an increase in the proportion of telogen hair follicles. Remaining hairs may be smaller in diameter and grow more slowly. One hypothesis suggests that melanocyte loss and lack of melanosomal transfer may increase oxidative stress level in highly metabolic hair follicle keratinocytes, affecting their function and viability.72 The process termed balding results primarily from the androgen-dependent conversion of the relatively dark, thick, terminal scalp hairs to lightly pigmented short, fine, villous hairs similar to those on the ventral forearm. Women are affected less often and far less severely than men. However, in post-menopausal women, hair loss is also the result of decreased estrogen levels and estrogenandrogen ratio.73,74 Besides hair loss, almost 50 percent of women older than age 60 years display mild facial hirsutism, presumably attributable to the same hormonal changes as scalp hair loss. In susceptible women, testosterone and/or progestin derivatives that are present in some hormone replacement regimens may exacerbate these changes. CUTANEOUS GLANDS AND NERVES Eccrine glands decrease by approximately 15 percent in average number during adulthood in most body sites. Spontaneous sweating is further reduced by more than 70 percent in healthy older subjects as compared with younger controls, attributable primarily to a decreased output per gland, predisposing the elderly to heat stroke. Apocrine gland size and function also decrease with aging. Sebaceous gland size and number appear not to change with age, but there is an exponential decrease in sebum production in both men and women most likely due to a decrease in production of gonadal or adrenal androgens. Pacinian and Meissner's corpuscles, the cutaneous end organs responsible for pressure perception and light touch, progressively decrease to approximately one-third their initial average density between the second and ninth decades of life and display greater size variation and structural irregularities. Decreased sensory perception in old skin encompasses optimal stimulus for light touch, vibratory sensation, and corneal sensation; ability to discriminate two points; and spatial acuity. Cutaneous pain threshold increases up to 20 percent with advancing adult age, and compromised arteriolar constriction on changing position from supine to standing is reflective of decreased responsiveness of the sympathetic nervous system. |